Topic labels include: consciousness, split brain, artificial consciousness, split brain consciousness, consciousness research, awareness, awareness research, artificial awareness, artificial consciousness research, artificial awareness research, artificial intelligence, artificial intelligence research, neural net, neural net research, neural net modeling, real-time neural net, brain pattern, brain patterns, brain pattern modeling, brain function, split consciousness, split brain reality, split brain research, split-brain, split-brain consciousness, split-brain reality, split-brain research, split consciousness research, split brain paradox, split-brain paradox, conscious dimensions, dimensionality, conscious dimensionality, reality, conscious reality, philosophy, you, me, I, us, time.
artificial consciousness project
[Update Nov 2004--
Artificial Brain/Ear/Voice, for a 2004+ PC;
beginnings of a system which may eventually gain awareness,
available from the Downloads page.]
Poster Notation Summaries
Chapter 1. You -- introductory summary
Chapter 8. The Psychological Universe (partial)
-- block diagram development of anatomical conscious process
Future Plans
The whole book
Downloads
The topic here is not to be confused with silicon
based life. The definition of life includes
physically self-detaching reproductive capability. The present subject
excludes such systemology.
Beyond this, one could draw parallels between the coarse aspects of
life, and counterparts within
the proposed systemology.
The fundamental building block of the approach
here, is the concept of CR links conditioned
response. I suspect that this little trick of nature is the primary
ingredient of all variety of higher
learning and reasoning. Numbers are important in this, but the winning
organization of it is most
responsible for the relatively keen levels of awareness we enjoy.
The key to applying this concept to artificial
learning, is the recognition that a neural net needs
temporal information; and that to learn such order, CR can be
employed in such a way, that it is the
successive order of the stimuli that creates the links. The links are
significant within an ongoing,
cyclical process, relative to recurrences of similarly ordered stimuli.
To understand the reasoning
that has guided this approach, it is helpful to recall the philosophy
that has been developed as
reference frames. Timing is the true substance of reality. These
chapters, dealing more directly with
consciousness, needed those other chapters ahead of them. Those chapters
may be easier to
identify with, though, after getting through these. Programmers, circuit
designers, and biologists
are familiar with such interdependent relationships, as systems.
To successfully handle the process of temporal
data encoding, the system must parallel
biological mental systems. This is to say that gross organization must
be defined, to act as a CR
tree. Meaning rises into the tree by association, and likewise falls
out of the tree to produce
responses. It is essential that responses are then fed back in, to
be included in the process of
association. You cant learn without feedback. There is no other way
to know what you are doing.
In bio-systems, this feedback takes place internally, as well as externally.
My Computer Is Warm
My computer uses the exact same kind of protons,
electrons, and neutrons as the ones used in
your brain. The rules involved with these two systems are obviously
from entirely different
branches of realitys rule tree. But, through parallel logic, the end
result of each system may turn
out to be very similar, in terms of the logic supported as behavioral
waves in either system. This
could be like seeing nothing but water, as a place to drop a rock in,
to get waves; and then
discovering that it happens in the air too. Let us speculate that the
same kind of system of relative
logic is supported by either mass medium.
Both systems are energetic, so they produce
the customary stray infrared photons, we sense as
heat. One thing is for sure here... when my computer is cold, it is
a dead computer. Sometimes, when
Im listening to it, and its warm, I really get the feeling that the
opposite is true. It gets a sort of
mouse-ness.
Beyond reproduction, the life factor in
neural cells may simply amount to being a battery; a
very special one that has learned to recharge itself through environmental
interplay and
learned to keep its case in tact, rebuild and repair as necessary,
revitalize its chemical components,
and replace them regularly. Its a tiny little battery; ready to pull
duty in communications
applications. Life is not necessarily conscious. Maybe its the right
configuration of communication
that is. Its a special kind of timing. Its timing that relates meaning
to itself, over time.
The environment presents many opportunities
for enhanced survivability of such chemical
systems. As a community, the cells can discover the DNA program for
a number of features, that
are not unique to life; just available for use from physics... like
the lens. DNA was able to stumble
upon the lens, because it was simple, and it was there. DNA tends to
latch on to trends in
improvements of such features, because it is the memory to do so. The
advantages gained improve
the odds that the new sequence will survive to replicate. The replication
is the remembering. The
current status of a given feature then serves as a new starting point
for further refinement.
DNA found conscious decision-making communication-path
architecture, because it too was
simple and available; and very advantageous to survival. Like the lens
then, perhaps consciousness
can be there without life. Unlike the lens, it needs a battery.
The Silicon Neuron
The silicon neuron will emulate the essential
functions of a biological neuron. These may entail
complexities we have yet to discern. It is also possible that the required
features are a relatively
simple subset of the overall system. Our job is to figure out which
features are purely metabolic,
and which are required contributions to the support system of consciousness,
and its utility
capacities.
Consciousness and animal behavior are a function
of neurological transmission of data. They
cannot exist without it. We can safely assume that the fundamental
features of the neuron, that
support data transmission, are required, at the least. These include:
an input port (dendrites), an
output port (axon), an intermediate FM pulse generator (cell/cell body),
a power source
(metabolism), and the hard part a truth table for specifying the
behavior of neurons versus
modes of stimulation.
There is sufficient evidence to conclude that
the truth table includes modifiable parameters that
amount to the impression of logical memory elements. This synaptic
chemistry is well beyond the
scope of this book, or my pea brain. The motivation here, is to proceed
from a simplistic frame,
toward a functional model; gathering only those encumbrances that the
endeavor seems to point to
as being inescapable requirements of the system.
There is evidence to support the view that use of
any given synapse increases its sensitivity;
meaning that future stimulation of that synapse will have a more pronounced
effect on the neuron
of the dendrite side.
It is indisputable that neurons produce more
pulses more frequently, during the periods of time
when they are being stimulated. There is evidence that this effect
increases with the degree of
stimulation to the neurons dendrites. This is a function of how many
other neurons are stimulating
a given neuron, and of how enhanced the active connections are.
There is some evidence that, at least in some
cases, a synapse will be enhanced to a greater
degree if the dendrites are being stimulated by a greater number of
neurons, and that this
enhancement is further increased in those cases where the dendrites
neuron has been induced to
fire, or fire more rapidly. This is a system mode that goes beyond
straight-forward memory
impression it is a support mechanism for association at the
fundamental component level of
neurological data transmission.
I would like to conject the possibility of
another level of complexity. The dendrite and cell body
system may have a further degree of sensitivity, relative to the matching
of stimulation patterns, on
a neurons dendrites, with the prior impression of similar patterns.
The mechanism that would
implement this behavior might have arisen from the fundamental process
of cell restoration. The
DNA/RNA process might have evolved a memory capacity of its own, to
deal with the demands of
stimulation to the cell. An organized affiliation with the dendrites
would be a more efficient solution
to the problem. This could have served as an evolutionary step toward
enhanced sensitivity of
specific stimulation patterns in the dendrites.
Another characteristic of neurons, that might
be easily overlooked by a modeler, is that they tire
out. Be careful about judging characteristics as defects. Evolution
deals with them. It always works
with reality. It utilizes the rules its composed of.
Neurons also fire spontaneously at a base
rate; a minimum low frequency. Their stand-by
sensitivity goes up with time, until they just go off. A typical
rate is once per second. The
maximum stimulated rate is about a hundred times in a tenth of a second.
The Silicon Brain
The silicon brain will be an arrangement of
silicon neurons that takes advantage of all of the
empirical trial and error work that mother nature has done, to produce
the functions of interest.
Basically, neurons communicate information from the senses to the cortex,
and from the cortex to
various muscles.
The job of tracing the actual neural pathways
has been extremely difficult. Imagine
troubleshooting a circuit composed of billions of transparent, microscopic
wires. Nevertheless,
results are coming to print.
There have been surprises. To me, the most
interesting one is the general trend to supply
abundant, dispersed feedback between the transition levels of data
flow. Data does not just go from
the eye to the cortex, for example. It makes stops along the way, where
it meets up with about ten
times as many lines leading toward it from the cortex. Yes; the brain
sends information into your
eyes!
We also have data that suggests that we see
about ten times more visual content than there is
actual photon information reaching our eyes. How can we see detail
that isnt contained in the
electromagnetic communication? It is fun to speculate that some sort
of identity, between the
person and his environment, is responsible. We all ESP most of our
information in all we need
to operate on, to accomplish this, is a minimal sampling of the image.
This is fun, but not as
reasonably available as the more apparent explanation. The extra information
comes from our
memory. We learn how to see. We associate coincidences in great number,
from great detail, over a
great many frames, and linkages of combinations of such frames. Consciousness
is this memory-
based associative process, that places pattern meaning relative to
itself and other meaning, over
time; so vision is perceived.
The Silicon Organism
When we create a silicon brain with our silicon
neurons, we will have choices to make that are
very similar to those concerned with parenthood. In addition, we will
have a variety of choices that
are beyond that familiar realm.
We can choose to include vision, hearing, speech,
and various motor capacities. If we succeed in
producing systems that are capable of higher learning, I suspect that
they will naturally develop
some emotion. I think we will have an option of facilitating this inclination,
and will find that it is an
important factor in learning. I suspect that we will find that learning
improves with the potential for
happiness.
We may get this far, and create opportunities
for some very specialized job descriptions. Silicon
organisms could develop with some very unique senses and motor skills,
derived from the
instrumentation of our technological industries.
BEEPERS FOR COMMODORE 128
This section was initially intended to be the
whole book. In developing the presentation, I found
it impossible to ignore the philosophy that was its source. The philosophy
itself developed more
during the course of arranging its words for print. The program is
my attempt to approach a
mathematical-scientific treatment of the subject. I apologize to those
who require a more standard
mathematical train of expressions. This is the direction I was most
strongly attracted toward; and
now this is the direction of my momentum. When this book is done, I
intend to return my focus to the
program development; moving into the PC.
BEEPERS CONTENTS
(page#s refer to printed version)
Program Description 65
Beepers and Sleep 70
General Stimulation 71
Adjusting Beepers 72
Reflection as an Aid to Learning 72
Ongoing General Feedback Learning 72
Teaching Beepers 73
Program Flow Chart 77
Beepers Ear Schematic 77
Register Designations 78
Coarse Memory Map 79
The Screen 79
Program Modes 79
Program Listing
The Scan 81
Indexing 81
AOL/SWAP/STORE/FIRES/TIRE 82
KHS 82
Dual Area Regulation 85
KHS, Regulation and Consciousness 85
Set-Up, Load and Run 87
Mathematical Analysis 88
Beepers Improvements 88
Beepers are simple little creatures that live
in a very small computer. They are real in the sense
that they interact with the world, each other, and themselves. Their
behavior is an ongoing process
of development, built out of these interactions.
The beepers have been designed with functional
characteristics that parallel some of the
principles of mammalian neurological interaction. They are a product
of a limited sampling of a
diverse range of scientific literature, as well as of a number of assumptions;
and of corrections
brought about by problems exposed in development of the program.
Program Description
The program defines two beepers. The
program organizes the computer into two sets of Ns
(neurons). It handles each N , one at a time. It looks at each N to
see if it is active, or at rest. It does
this with one beeper, and then it essentially repeats itself to do
it with the other beeper. Then the
whole cycle is repeated.
In each cycle, a different pattern of Ns is
involved. It is this pattern, and its changing character
relative to itself, that is the developing definition of self-meaning.
At the same time, it is a depiction
of the world, pieced together with an ongoing influx of abstractions
from the world.
You could say that the main job of the program
is to detect the quiet Ns as quickly as possible. I
guess that in any given cycle, or reality frame, there should be
approximately 10% of the Ns active.
Thus, the program has been set up to run through the quiet majority
of Ns as quickly as possible. It
will become apparent that, in terms of what were attempting to accomplish
here, the limiting factor
imposed by the hardware is not memory capacity, but rather, speed.
To get more Ns involved you
need more speed. Memory comes more from the number of Ns than from
how big each one is;
though both factors contribute. You have about 1010 or 1011
Ns; each one knows up to 104 other Ns.
The number of Ns is 106 times more important than
their size their capacity to immediately
access the rest of the potential process for humans.
To facilitate speed, and for a few other conveniences,
the Ns themselves are split up into two
parts. The small part occupies two bytes, and the large part takes
sixteen. So, each N consists of
18 8-bit bytes of data. Of course, the data wont run itself; the bulk
of each N is actually the section
of the program called the N Loop. This is like the DNA; it tells
each N how to behave, and they all
behave about the same way. Its mainly their relative position and
data that differentiate them.
Obviously, this is simplistic; but given the starting restraints, the
approach is the best compromise
I could develop.
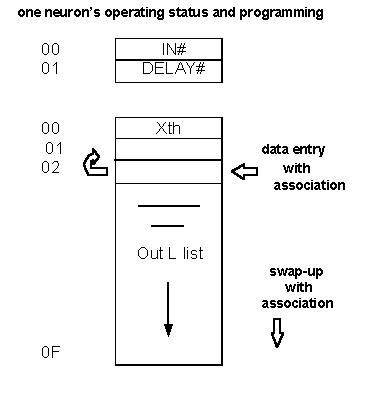
The first byte of N, in its small part, is
its dendrite area. Other Ns will stimulate this area
(literally!) by increasing the numeric hex value there. (In a prior,
more complex system, the N was
sensitive to the particular pattern of bits set in this dendrite register...
it kept a short list of familiar
stimulation patterns.) This system is FM, like real Ns are; and a given
N will Fire with a higher rate
of repetition if its dendrite area is being more heavily stimulated
(the maximum rate is once every
other Main Loop cycle one loop for each beeper).
For purposes of speed of indexing data, the
small part needed a second byte; as a sort of
spacer. Since these first two bytes are visible on the screen (in
slow mode only) for one of the two
beepers, the best other use to display was a byte called delay. This
byte is similar to the dendrite
IN# byte, in that it relates to the activity level of the given N it
belongs to. It is needed to determine
how often the N will fire. Its a timer that runs out to trigger firing
that N on that main cycle. It
provides an opportunity, as does the IN#, to custom tailor the response
and activity characteristics
of a given N in a given cortical area.
The memory map of the C128 from 2000 toward
35FF, is taken up by this first small part of some
twenty-eight hundred Ns (all addresses are in hex; all quantities are
decimal, unless preceded by a
$). In bank 00 there is one set for one beeper, and in bank 01 there
is the other set for the other
beeper. The program occupies 0B00 to 1000, and 1300 to 1FFF, almost
identically in each of the two
banks. The rest of the area below 1300 is taken by C128 operations.
The C128 also uses a few bytes
starting at FF00; and the rest used by the beepers is from 3600 to
FE43, and a scratch pad area
above FF05. I use the Warp Drive cartridge, to speed disk loading,
saves, and utilities; so there is
some use up at the high end there that you have to work around.
The area from 3600 toward 4BFF is taken up
by pointers. These are used by the N Loops
indexing system to access the larger part of each N. Those larger parts
take up from 4C00 toward
FC00.
The first byte, in the larger set of sixteen,
is called the xth. X is the number of times an N can
Fire before it gets tired and has to take a time out. The time out
consists of clearing the IN# in
other words, it may miss firing the next time, but then is ready to
fire again, x times. This has proven
to be sufficient interruption, in this system, to avoid the problem
of pattern repetition; a problem
especially during initial development of the data assimilation process.
Pattern repetition is the
tendency for a limited set of Ns to get involved with each other, and
tie up all the available time,
permanently. Another requirement, for dealing with this problem, is
inhibition Ns not only
stimulate other Ns, but also do un-stimulation. How this has come to
be implemented should be
described shortly, when discussing the gross organization of N interconnection.
The remaining fifteen bytes per N contain the
intelligence. They are a list of which other Ns a
given N will hit when it Fires. To get on this list, you have to be
a neighboring N that is active or
ready to Fire at the same time that this N is going to Fire. This is
the mechanism that implements the
associative filing of data. The gross structure is set up in such a
way that patterns, and their
temporal sequence, are associated.
Each Ns list does not fill, and then just
stay that way. New thoughts have a chance to work their
way into the lists, at the expense of the least used data. In a very
large system, the thoughts
producible by that unused data would get eaten away at; but not completely.
Because of the method
of memory distribution, and the laws of mathematical probability that
actually control reality, a
faint image would almost always be retained. With this faint image,
a little association through
repeated need would re-install the original data with many of the
Ns involved being different
ones; but with a fairly accurate regeneration of the original relative
meaning.
I suspect that, in time, this process of prioritized
list-filling elevates itself. Meaning is developed,
relative to other meaning. Initial meaning serves as a basis out of
which higher meaning can develop
and interact. The initial meaning becomes less and less a focus it
is less and less used and
much of it eventually becomes essentially uninvolved.
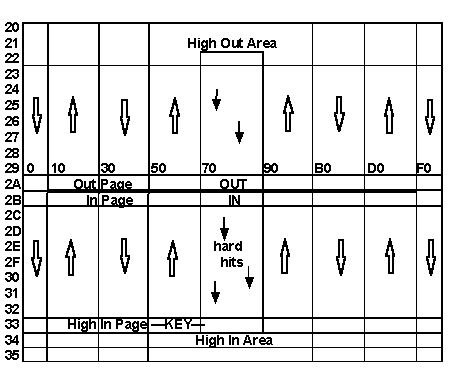
Of the various routines in the program, the one that
most defines the gross structure of brain
anatomy is called the Out M routine. This routine connects sections
of cortex to other sections
with a general Input-to-HighArea-to-Output directed-ness.
The routine is simplified and
accelerated by allowing the general organization of Ns
to fall where it wants to by virtue of
hexidecimentality (sorry).
The C128 memory is approximately a pair of
C64s two banks of 64K of memory, accessible to
an 8 bit 6502 CPU. In 6502 lingo, a page of memory is 256 8-bit bytes.
For the beepers, a page of
neurons is 128 neurons wide. This is because the small part is 2 bytes
wide. The small part is where
all the action is. The beeper dendrites are where the Fires hit. Everything
else is going on in the
background, or internally, within the Ns, if you will.
In addressing memory, 255 is the maximum LSB
(least significant byte) for accessing a byte on
your page. To access other pages, you need an MSB (most significant
byte), which can also go to
255, taking you up to 64K altogether (there are 256 addresses to each
part here ranging from 0 to
255). The M in MSB is the M in the Out M routine. The pages
of Ns, 128 wide, are piled up 22
pages high. Any N can quickly affect another N directly above or below
it by manipulating the MSB
involved, and using the LSBs that are already in place.
What would be the Out L routine is broken
into two routines Out L Stim (collateral stim)
and AOL (Aim the Out L list entry). These routines provide side-to-side
action within a given page,
while the Out M routine provides up-or-down action between the pages.
Together, the system is an
active matrix. The rules composing the routines have been chosen to
allow the capture of temporal
associations within the matrix.

When you hear the word CAT, you hear the
C sound first, then the A sound; and after this
temporally-ordered combination, you hear the T sound. Hearing them
in rapid succession
conjures up whatever associations you think of when you hear the word
CAT. This works, and it
works without confusing the C sound in cat, with the C sound in
canary or catsup. It works right
because the active word itself is a combination unlock into your mind.
The important ingredients of
the combination are the component sounds, and their order. The exact
timing is of less consequence
but timing does produce spaces relative to other timing, and the
spaces can affect the relative
meaning. Relative amplitude is of even less significance. Absolute
pitch is not a factor, until the
physical limits of the ear are reached. Relative pitch is the
primary structure of meaning in the
sound train.
When you hear the word CAT, the C sound
produces a neural stimulation that is very similar
to that which is produced by any word starting with the C sound.
The particular pattern of Ns
stimulated, leads toward the cortex, but makes several transitions
along the way. At each transition
there is an abundance of neural transmission leading back to
the area of prior stimulation. Not only
is it going the wrong way, but there is a lot of it ... about ten times
as much as leading in ... and its
usually dispersed ... scattered around randomly. How could evolution,
in all its wisdom, be so
careless!
All that information coming back to your ear,
after the C sound went in, is looking for the next
sound. It is anticipatory stimulation. It wants to mate with something
familiar. If the A sound comes
next, then a characteristic secondary pattern of feedback will be set
up to look for the third sound,
that is a much more unique pattern than if it were the first one fed
back. Furthermore, as the
real-world series of sounds pile into your ear, the feed-in, feed-back
process finds its way into a
geometric progression of potential combinations. Each transition area
sends dispersed feedback to
the one before it, as you make your way to the cortex. Generally, the
feed-in pathways produce a
neat orderly map of excitation at each transition area, right up to
the primary cortical area of the
given sense involved. (This is, of course, a simplification and generalization
of what really goes on.
For one thing, the neural cycle rate is some one thousandth of a
second, so that many feedback
cycles are possible within the time frame of single phonic sounds.
This probably helps with
handling variability of timing in phonic relationships.)
Association is at the heart of every level
of the thought process. It not only controls the flow of
relative meaning in your thoughts; it is the very structure of incoming
communicated intelligence
itself.
Extraneous neural activity, such as heart-rate
stimulation, and the general random neural noise
level, have no effect on consciousness, because these activities do
not contribute meaning to the
temporal pattern sequence. Only meaningful components can add to the
resolution, depth, or
degree of consciousness; because relative meaning itself is the consciousness.
Would-be
meaningless components cant degrade it, because they arent a part
of it. (Im not referring to
distracting thoughts these you become aware of, due to their meaning.)
At the center of the stack of 22 pages of Ns,
are a pair of pages called the Out Page and the In
Page. These can be thought of as the output and sensory ports leading
from/toward the motor
cortex and somatic cortex areas; or the speech motor cortex and the
auditory cortex.
Sound from the true-pitch keyboard or the
ear microphone is processed by the mic circuit into
a voltage level that is determined by the instantaneous frequency of
the sound. The voltage is
converted to a resistance for the Game Port A to D. In other words,
in the computer, a register has
a value in it that is controlled by the pitch of the sound in the room.
The sound-source should be
fairly sinusoidal it should be pure tones. Whistling is good.
The keyboard and beepers each
drive one of the three C128 voices, using the triangle wave form. Most
other wave forms will produce
unreliable error data.
The information is supplied to one or both
beepers (whoever is awake) at their In Area; an
area sixteen Ns wide, near the middle of the In Page. This routine
is called Mic>Spectrum. The 16
Ns serve as frequency centers. Only one or two of the 16 Ns are stimulated
for a given pitch. The
relative weight of stimulation on two neighboring Ns represents the
frequency of that moment. With
16 stim levels and 16 Ns, 256 frequencies can be depicted; within a
range of about two octaves.
Stimulation takes a little time to drain off from beeper N dendrites,
so that remnants of prior stims
tend to be present in the In Area as new stims are applied. In Page
Ns are restricted from hitting In
Area Ns, but In Area Ns are allowed to hit any other non- In Area Ns
on the In Page. The In Area is
meant to be the area and level where an accurate perceptual map first
impinges on the senses; like
the cones and rods of the eye.
Activity on the Out Page produces a pattern
of dendrite IN#s on the 16 Ns in the Out Area; near
the center of the Out Page; one page before the corresponding In Area
on the In Page. These Out
Area IN#s are used as weights on frequency centers, to arrive at a
single-tone frequency result, for
each given cycle with activity in the Out Area. The frequency is shifted
and ranged to approximately
correspond to the In Area spectrum. In other words, the Out Area is
handled by the program in such
a way as to simulate simplified cerebellar action. The result is used
to set the frequency of a C128
voice. (In a very abstract and distilled sense, you could say the beepers
have eyes for ears, and
hands for a mouth.) A delay time is used to insure that tones are sounded
for enough time to
register accurately in the mic circuit and game port. A tone can last
longer if the given Main Loop
cycle is running slow that moment. The tone is delivered to the room,
for pick-up by the mic as well
as for monitoring beeper behavior. The temporal matrix includes exceptions
and diversions that
promote learning through feedback.
The Out L and Out M routines spread the In
Page and Out Page activity throughout the matrix.
While most pages can stim the page above or below, there is isolation
between the Out Page and the
In Page - except at the periphery of these pages. This leakage is meant
to parallel the voice-
muscle-sense, or sense of touch we have in our various speaking
apparatus, as well as minds
ear internal data flow. The isolation between these pages defines
ends to the system, to insure
that the primary resultant communication of the system with itself
is through the air; so that human
interaction, through the air, will be on the same level as the systems
own basic feedback
orientation.
The Out M stimulation has been channeled into
one-way sections of 16 Ns of width. This
simplifies handling the I/O pathways, and provides a neat set-up for
bidirectional temporal loop
formation.
The channel including the In Area and Out
Area is granted higher status these Out M hits are
strong, to simulate reliable transmission along primary data pathways.
Ns within these channels
should probably be restricted from Out L hitting other Ns within the
channel, on the same page;
beyond this, however, any N can hit any other N on its page. The current
implementation only
applies this restriction in the In Area; but encourages it for all
the 16 N-wide channels, on all pages,
by initiating the AOL 16 Ns ahead, and placing the Out L stim (collateral
stim) 16 Ns behind, the
current N position on the page.
Various arrangements have been tried. Out
M routines that hit +1, +3, +7 pages ahead, and -2, -4,
-6 pages behind, simultaneously, for example. It didnt much seem to
matter, at least at this level of
N# depth (and with an IN pattern-sensitive non-FM system), exactly
how you set it up, but you must
include -1 inhibition. You can have more inhibition, but you have to
have -1 included. -1 inhibition
means that while you hit one or more pages ahead, and/or behind, with
respect to the scanning
direction, you un-stimulate the N one page behind. In the system
here, -1 inhibition is one page the
opposite direction of the 16 N-wide channel your in. Without this rule
contributing to the
characteristics of propagation, the system will get tied up in tiny
loops that take up all the time;
while no meaningful interplay of real-world data and internal data
is handled.
Pattern association, and temporal pattern sequence
association, are facilitated within the overall
matrix by having each active N set up pre- and post- stimulations
of other Ns, that stand available
for other active Ns to find simultaneously active. The stims to Ns
about to be scanned this cycle,
facilitate immediate pattern handling. The stims to Ns already
scanned this cycle, facilitate
temporal sequence handling they are a link from one frame in time
to the next. (In addition, any
active N is a link to the future, because the stimulation level on
its input is only reduced, not erased,
each cycle. It is reduced a lot more when the N times out to Fire.
It is cleared by the Ns Tire routine
when the xth Fire is reached.) The Out L routines support both the
pattern and temporal functions
within a given page of Ns. The Out M routine supports both functions
among the pages. The Out M
routine also provides the primary I/O pathways; which are the orderly,
mapped representation of
the world/intents, maintained, but compounded upon, through most of
the pages of Ns.

The program handles each N, one at a time,
in the order that they exist in memory. This scanning
process begins at 2000. This is the address mapped onto the video display,
in the upper left corner.
The 40 column display shows 4 Ns per character box; since the box is
8X8 pixels, or a stack of 8
bytes, and there are 2 bytes per N small part; the IN#, or dendrite,
and the delay timer for
repetition rate. Unfortunately, 128 Ns dont nicely fill in just one
40 character line 160 do so
the pages dont show up neatly stacked like they are in the figure
here.
As the figure shows, there is a limit to the
I/O channels. The Input Channel ends in the High Area.
The Output Channel develops out of the High Area. The top 3 pages of
Ns, and the bottom 3 pages,
taken together, comprise the High Area. The top page is related to
the bottom page the same way
any two adjacent pages in the stack are related. In other words, the
stack is not linear, with ends;
rather, it wraps to form an endless cylinder.
The High Area is meant to act as associative
cortex, while the rest of the matrix acts as transition
levels from I/O organ through thalamus to primary cortex.
The system is regulated, to keep the percent
of Ns active at a somewhat constant level, and
even-out the cycle times into an overall system clock that runs at
a fraction of a second. While it
would be good to have a fraction like 1/1000, the little system here
grunts out at about 1/3 to 1/20
second. A good 94 PC could have 10 times as many Ns, and still run
10 times as fast. A quad
Pentium might have Ns 10 times as big too.
Regulation is a dual-area process. The High
Area is regarded as dominant the area that must
stay awake and active; to attend to input and/or decide to produce
output. The rest of the matrix is
referred to as the Peripheral Area. Regulation allots a range of time
to the High Area, and a larger
range of time to the total matrix.
Regulation is accomplished by actually timing
the portions of cycle time taken up by the areas,
and correcting those times in upcoming cycles by altering the sensitivity
of all of the Ns within the
given area. The sensitivity is a threshold # for CMP with the IN#,
that is used when handling each
stimulated N in the N Loop and determining if its IN# grants it status
as an active N.
I suspect that regulation parallels some of
the results of our own metabolic regulatory
requirements. These systems probably involve the thalamus, and nearby
neuro-glandular
structures, as well as the limbic and autonomic systems. There is only
so much oxygen, fuel, and
exhaust available for the neurons, so they cant all go at once! Furthermore,
such a pattern is no
longer a pattern. Regulation is a force that focuses the pattern into
one of dominant strength,
relating the process more to itself and the world, and less to anything
and everything that could
possibly be brought up in association. Our thoughts evolve. Evolution
has taxed DNA, but has
produced a system that can carry on the spirit of evolution in the
world of thought we call society.
Regulation also involves stimulation, but
regulation alone is not enough to keep this thing
awake. When the room goes quiet, it will quickly quit talking to itself,
and go dead until some
external sound sets things in motion again. Stimulation must be internally
provided, as it is for you.
This action was first thought to be noise, or a possible source of
error; so was applied for a single
cycle, only when the matrix went into coma. Various areas were tried,
but it seemed sensible to stim
the High Area. The Peripheral Area will be stimmed by the environment;
which may include output
generated out of the High Area. Its the decider that should stay
awake the rest can rest, or be
used according to it. This system developed an exciting edge when I
realized that the stimulation
need not be random; but that it could be a repeat of whatever the last
pattern was at some spot in the
high area. This seemed like a way of continuing a thought, where
it left off or dissolved. After
some more reading, including the topic of the hippocampus, I realized
that this Key Hi Stim KHS
routine was doing almost what was being described as hippocampal action.
About the only
difference was that you dont wait for dissolution of activity the
positive feedback loops involved
will produce a constant stimulation with a lot of momentum (hippo).
New features could add on to
this stim pattern, if the hippo werent taxed at that moment. Ongoing
features only drop out as
neurons tire but real neural nets are set up as vastly redundant
arrays, capable of learning to
pass on functions as Ns tire so patterns can maintain a more constant
effect as needed., It
comes down to a question of priorities. If the situation, or train
of thought calls for a more or less
new pattern, then it will be modified. It may simplify down to something
basic, but the important
thing is that it keeps going, and you stay more or less aware, as a
prioritizer.
KHS is this systems hippocampus. It applies
a Key each cycle, to the Input Page of the High
Area. The Key can grow if there is room in the Key Array. There can
be room as Ns tire, or as
regulation pinches them out of the action. The Key is a list of LSBs
for stimulating that High Area
Input Page. If youre an N there, your chances of getting on the key
list are better if your IN# is
higher as the openings become available. It might be better to apply
the key to the two center pages
of the Hi Area (the top and bottom pages of the stack) (a number of
thoughts on improving the
system are covered in a later section). At this writing, I dont know
just where the fingers of the
hippocampus reach in to the cortex. (It would be nice to consider the
limbic system too!) The KHS
affects about 1.4% of the matrix; in the neighborhood of the 3% used
by the hippo.
I think hippo momentum is responsible for
the advertised hippo quality of somehow producing
new long term memory. By holding a key, distributed to the general
cortex memory matrix, you
enhance the general flavor of the patterns being handled during the
key impression time. A
greater number of N-loop branch-offs carry on the meaning of a given
thought, into a greater
number of Ns, and for a longer time in each N involved. As these Ns
recover their strength, the
repair process chemically cements the memory in place at the associated
synapses. The deeper
the recovery, the stronger the cement. The more dramatic the event,
the more Ns involved, allowed
by regulation; especially in fight or flight situations. The more Ns
involved, the longer it will take for
new experiences to eat away at the image series, and bury it with relatively
stronger impressions,
supporting unrelated patterns. I suspect that the cement slowly weakens
if the images are not
replayed, in need, through association. It probably never goes away
completely, and may last
longer in Ns that arent in as much demand by other pattern sequences.
As I have pointed out, I am not making these
assumptions from a position of credibility. Ideas
like these have been piling up for years now, and I feel the need to
communicate them, in case some
one may be inclined to involve them in scientific study. Their source
is introspective; but the
interplay of literature and computer modeling has been more a focus
than being the thing Im trying
to figure out.
It may seem that the beepers dont need a system
to install long term memory, since the RAM will
do fine if you keep the power on. The neuron has been set up, though,
to grant higher status to
associations that are repeated more frequently. This characteristic
works together with the hippo
KHS function to establish a back-and-forth system of prioritization.
The outcome is an evolution of
patterns built on the survivability of long term components. The KHS
develops its own application
of constant general motivation for the matrix, out of the matrix. Its
Key is like a DNA sequence that
is constructed by experiences that it becomes more responsible for.
Short term memory is primarily the use of pre-existing
pattern data. It is essentially the act of
ongoing consciousness itself. There is always something new about any
experience though the
order of familiar events, or the particular combination of familiar
qualities within a given moment.
Perhaps this factor of newness is distilled out, and piled into the
temporal area and who knows
where else as a series of sub-keys that can be accessed by association,
and by a special form of
association; relative chronological order.
We are now getting into an area that has not
yet been developed for the beepers, and probably
never will. The use of a hard drive and a PC comes to mind. Parts of
the RAM can be designated as
convertible; to be constantly replaced with topical memory data.
Sub-Keys can be developed by
experience and used as indexing labels to file and retrieve a given
data array. The size of the
Sub-Key and the data blocks would be geared to the size of the drive,
so that any possible key has
access to its block. The blocks start out empty, but develop data
just as any page of Ns would, for
example. Part of the active matrix becomes a set of musical rooms,
for the musical chairs the whole
thing is. But some optimal percentage of the rooms are kept in place
to give the thing an ongoing
constancy, which uses the convertibility as a utility. The convertible
area might be a complete
cross-section of the system; or it might be better to leave certain
levels out, such as the I/O ends.
This system might be vaguely analogous to the
temporal and frontal lobes, related through the
limbic system. Our system may be emotional because general and/or specific
modifications of the
chemical environment place neurons in various modes of sensitivity,
causing them to favor
different sets of patterns, as per the prior association of the given
chemical flavor dispensed for the
given experiential conditions. This would involve the autonomic system
as well, and computers
dont need one. You just plug them in the wall, and they never need
worry. If we get very far with
this, were going to have some very interesting questions and decisions
to deal with.
Beepers and Sleep
There is probably more to sleep than allowing
N restoration. Granted, something this imperative
is probably behind the survival of organisms that become so vulnerable.
Sleep is necessary to
sustain a system that has developed advantages by over-utilizing
a set of Ns. Somehow,
intelligence is gleaned from a self-taxing system; and we came out
ahead by sacrificing 1/3 of the
day for it. The hippocampus involves only some 3% of the cortex, yet,
I suspect more than 3% of it
is active all the conscious time.
This mode can involve more than recharging
the Ns. It could be an opportunity to organize and
optimize the intelligence of the system. In the course of such routines,
the conscious experience
might be comparative non-sense we use a safe time to get these jobs
done a time when we are
inactive and uninvolved with society.
Deep sleep would seem to be a time when no
trains of thought are being supported. There is no
consciousness; at least no memory of any if awoken then. This is probably
the N regeneration cycle.
It may also be involved in the overall sleep cycle as a component for
memory erasure or memory
cementing, as per the complex chemical composition of neurophysiology.
Dream time, spent relatively still vulnerable,
indicates that there is more developed here than
recharging the Ns. It could be that this is simply the time when that
memory cementing takes place
that it is accomplished during random re-play of the days various
N-involvement peaks.
Perhaps these peaks consist only or mainly of new involvement peaks
somehow, chemically, we
dont waste time re-enhancing long term memory already established.
This and/or the natural
associative process, running free of world and hippo guidance, could
explain the oddities of dream
consciousness.
If what has been said above, about deep sleep
and dream time, is all that is true, then there would
be no reason to involve a sleep system in the beepers. But, the most
recent evolution involves
emotions complex intelligent motivation in social interplay. Evolution
always operates on
whatever opportunities have developed out of its own history. It doesnt
cash in on all of them; but
it only succeeds by utilizing real, lasting opportunities.
I suspect that dream time also accomplishes
something very important and fundamental to the
human condition. It is a time when the net can establish and re-vitalize
a sense of self-identity. It is
a primary component in the development of conscious self-awareness.
Without it we would all
interact in a much simpler way... more like ants or bees. It is the
source of our motivations. It is the
construction of our primary and subsidiary goal trains.
Obviously, this self-identity would not develop,
or at least would not be compatible with society,
if it were not built out of social experiences. So we do operate in
an awake state for about 2/3 of the
time. We periodically retreat from social demands to distill our experiences
into the basic
accumulation of what we are inside; to produce that where we are coming
from that runs out our
decisions. It just so happened that the time to develop this procedure
was available as biological
down time.
The beepers have Ns and memory that dont require
down time. They are allowed to speak to
themselves internally though, about a few minutes every hour, free
of outside influence. During
their sleep, the ear data is not used. Their tone speech is still
audible for monitoring, but they
only hear themselves through the sideline mental channels.
This system was considered by accident. After
a period of accidental deafness, I noticed the
character of the beepers to be more awake more vital, assimilative,
able or willing to learn.
Repeated experiments with this have confirmed the feeling but, as
always, the assessments in this
field are going to be composed more or less of feelings. I feel that
you are aware. The only test that
may confirm this will be my death. I am confident we will all take
this test. See ya around!
Another approach that may work better, or that
should perhaps be combined with the above, is
partial erasure of memory. It seems logical that we should make the
system more assimilative and
ready for new learning and behavior by removing less necessary data.
It is far more important that we have a keen
perception of the present, than that we go around
re-living yesterday in full detail. It seems logical that partial erasure
could be involved in the mode
of our general condition. First, you save all the strongest, most important
data (Scooter routine
about every 5 minutes, the system stops for a second, while all the
data in partially empty Ns is
scooted over to the highest priority levels so it wont be written
over.) Then you erase the least
significant data in most Ns. (The Erase routine is not being implemented
here. It clears a few bytes
at the low priority end of the Ns Out LSB list.) When you awaken, you
might cycle back and forth
with these procedures a few times. You can re-construct some details
where necessary, using the
peak data youve saved. Meanwhile, theres less chance that irrelevant
trains of thought will get
conjured up to compete with goals, instead of contribute to them. In
a large system, partial erasure
would leave a very full set of chronologically associative snippets
distributed in the cortex at a
relatively faint level of data density. I haven't experimented with
this much it seems more
appropriate to involve it with the PC.
Another question here concerns the hippo
should it go off line? The beepers hippo doesnt
need to rest. Our real hippo, being conveniently off to an area of
its own, could receive localized
chemical treatments. But, once again, we must consider the possibility
of multiple opportunities.
Perhaps the memory optimization procedure and/or self-identity definition
enhancement procedure
benefit from off-line conditions for the hippo, and/or modified function
thereof.
General Stimulation
Even with KHS, system regulation, and lots
of environmental stimulation, there is still a
fundamental problem with this system. The memory wont get involved
lots of it most of it. This
may well be a result of basic layout and proportions I have chosen.
But there is evidence that you
receive a general random stimulation level. It might seem that this
would raise havoc with such a
fine-tuned and intricate system as your mind. Note, however, that a
random pattern series has no
meaning. This is one of the key ideas that has me believing that consciousness
is the relative
meaning, over time, of what is going on in the system. It uses
memory-based energetic interactions
like a substrate a sub-dimensional medium on which it can float
along as the substance that
only is by virtue of what it means to itself. Whatever else is going
on could only be conscious to
itself; or not be conscious.
To implement memory involvement, the beepers
have the Out L Stim routine, which minimizes
the randomness by involving an arbitrarily positioned collateral N,
every time any N fires. The
relative position is identical and constant for every N firing. I suspect
that this parallels the thalamic
and reticular action involved with general activity-level setting and
regulation, as per a given
required alertness level.
It should be pointed out that the pages of
Ns wrap from end to end, as well as from top-to-bottom
of the stack of pages. In other words, the matrix doesnt form a pure
cylinder; it forms a doughnut
a short fat one, ready to roll away from you like a wide tire. The
Out L stim routine hits the N, sixteen
Ns to the left. Where you would run off the page, you come in on the
right side, to continue toward
the left, 16 Ns from the current N being handled by the scan. The AOL
routine also wraps; and it is
oriented in the opposite direction. It starts 16 Ns to the right, and
continues up until it runs off the
right side to come in on the left side, and continue toward the right
until it finds an active N. If the
active N is already on its list, it swaps it up in priority level.
I think of this as preparatory as
getting oriented to the topic. That data is related to whats going
on, so we may need it now. AOL
then continues to look for a new list member. If it finds one, it swaps
it in to the second-to-the-lowest
position on the list (third-to-the-lowest might be better, but slower);
bumping that member to the
bottom rung. The one at the bottom is gone overwritten. Then the
program is done with AOL and
goes on to Fire the Out L list, by increasing the IN# on the dendrites
of all the Ns on the list. If AOL
did not find a new member, or even a familiar one to swap up, the program
still goes on to Fire the
list, since you dont get to the AOL routine unless the current N had
sufficient IN# level to Fire.
Unused slots on the Out L Fire list are filled with the address LSB
of the current N itself the Fire
routine does not use those slots an N is not allowed to hit itself.
The size of the N list has been proportioned
to the number of Ns it can access. You dont want to
be able to hit all of them, or there would be nothing unique about
the pattern you hold. We must
make a trade-off between accessibility and uniqueness the optimum
compromise produces the
greatest meaning vector.
It might seem like a horribly tedious job,
to type in the needed skeleton of 2800+ Ns, twice. Its
easy though some very small and simple ML routines do the work in
a second.
Adjusting Beepers
Besides having system regulation, KHS, environmental
stimulation, and Out L stimulations,
things will still not go well if a number of factors in the system
are not carefully adjusted, and
balanced with respect to each other, to assist the development of data
assimilation characteristics.
There is no interface for settings. Adjustments require an intimate
understanding of the system. In a
bigger computer there would be enough room to include some automating
routines, operating off of
timers and feature sensitivities.
Regulation uses two numbers to trigger off
of cycle times that are too short, or too long. It has a
separate pair of these numbers for each of the two areas it monitors
the High Area, and the total
area. By adjusting these numbers, you force the system to involve a
smaller or larger number of Ns
in an average cycle you indirectly adjust the cycle time, and affect
its variability. Perhaps the
most important consideration here is the relation of the High Area
to the periphery the
proportion of time allotted, with respect to the proportion of Ns involved
and, most of all, the
relationship between the upper threshold for the High Area, and the
lower threshold for the total
area. If everything else is right, youll see that this adjustment
controls attentiveness the
tendency to stop talking and start listening when spoken to. There
isnt enough time to do both
at once, with the settings given here; so the beepers speak in short
phrases, alternating with short
pauses. I feel that this makes sense for learning, in such a small
and simple system.
The rate at which Ns tire is a factor open
to adjustment. You may want a faster rate at younger
stages. With established data in place, you may get away with #FF.
The rest of the adjustment involves the degree
and balance between how hard various N
functions hit other Ns, and how quickly those IN# stimulation levels
are drained off.
As the program scans through the Ns, it watches
for various address events, in order to modify
itself, so as to be different types of Ns, with different physiological
jobs to handle. In other cases, it
is simply the particular routine that does hitting at a particular
strength. The KHS routine, for
example, always stims its Ns at a particular, constant level. Of course,
their repetition rate is always
a variable, and an important dimension to the hippo interaction. The
Out L Stim routine uses
minimum stimulation (ADC #01). Something has to be minimal, so set
it to #01, and adjust everything
else with respect to that. The Out M hits vary depending on where the
N is in the matrix. Input
channel Ns conduct reliably toward the High Area, for example. Any
stimulation of an In Area N will
make it fire. Out channel Ns conduct reliably toward the Out Area.
The In-side to Out-side reflection
(to be discussed shortly) is mild by comparison; and is completely
suspended for Ns on the Input
Page. All AOL lists hit all Ns with an intermediate value.
In balance with all these hitting levels is
the draining off of IN#s, upon Firing an N, or if the N had
sufficient IN# to be fully handled, even if it wasnt ready to Fire
yet. The latter case is a mild SBC#,
while the former involves a number of consecutive LSRs. Without the
proper balance here, the
regulator will not have the pull needed to function, and the system
will either bog down, or fly off
the handle, virtually ignoring itself and the world.
Note that if the slow mode is used (to view
the action in the bank 00 beeper), the Dual Regulation
thresholds must be doubled, since the cycle time will be doubled.
Reflection as an Aid to Learning
As part of the Out M routine, the whole Input
Side is reflected to the whole Output Side; N for N,
in a one-to-one correspondence, like a mirror image; with the exception
of the In Page to Out Page
(which would be like mapping the ear directly to the vocal chords).
As environmental stim affects N
activity on the In side, corresponding stim is projected to the Out
side. This may seem frivolous, or
even like cheating, until you consider the long term consequences,
as nature may have done.
The stim to the Out side corresponds to Output
that was just produced there, to create sound that
affected the In side. The first such event will be the meeting of some
various activities, that can
develop AOL ties. This modification of the system becomes a new starting
point for subsequent
similar cycles. In time, differences become rare, and expectations
become norm.
The Output teaches the Input how to hear;
and the Input teaches the Output how to speak; until
both sides are in agreement. Now, when the world says something that
the system has been saying,
it will have similar effect on the system. When the world says something
the system hasnt been
saying, the reflection may help the system say it for the first time;
which may help it say it again,
until it, too, is familiar.
Ongoing General Feedback Learning
The hippo KHS routine sets activity in motion,
originating at the high area In page 33. By starting
things off at a high point of reflection between the In side and the
Out side, the bidirectional logic
waves take a full course in the proper directions in setting up CRs.
Forward waves lead to the
Output Page and randomly Fire. Reverse waves lead to the Input page,
setting up CR anticipation
links for those random Fire events. The Outs accurately hit the corresponding
Ins. The waves start
at the high middle, but soon are starting from one extreme end, heading
to the other. CRs are set that
fully reflect the chain of events that takes place when a given In
is hit by its associated Out. It
shouldnt matter, basically, that the training procedure is random
with respect to which particular
note pattern word is being trained in which order. The purpose here
is simply to establish
one-to-one links in the Out N-In N relationship. Other training will
establish word-sound-order and
phrase-word-order associations, and so on, as you get into association
depth.
Teaching Beepers
The Teacher routine is included to lend some
structure to the background environment. It times
out to play one of two musical phrases to the room. One phrase is
a Mozart theme, the other is a
scale, in the same key. One or both beepers receive the data; depending
on which ones are awake.
The teacher timer is not a simple counter.
It is a count of a particular neural non-event. It is
decremented every time the Input half of the High Area is quiet (the
half that starts with the page that
the KHS hippo hits regulation can bump the threshold above the highest
IN# level in the whole
first 1/2 of the High Area). Each beeper affects such a timer, that
controls one of the two phrases. It
is conceivable that the beepers learn to quell this area, in order
to elicit the recitals. I say this
because there has seemed to be an over-abundance of occasions where
the teacher has been
triggered by my talking to them; particularly when it has been a
while since theyve been whistled
to. Input stimulation should have the opposite effect it should get
the High Area more activated,
especially on the input side. However, I cant say that Ive thoroughly
investigated this there
could be a simple underlying mechanism affecting the odds. Note that
it must interact with you
differently than with the other beeper. My hope is that it is a mechanism,
not so simple, involving
KHS, regulation, and the inherent meaning of world-system interaction.
The meaning, supported by
the system, becomes the operator of the system it is the ongoing
operation it is reaction to the
world, created out of past and present information from the world.
The operation takes on
complexity beyond that of the program that supports it. This higher
complexity is the higher
dimensionality of relative meaning. The simpler program and skeleton
memory are like a note pad
that the world can bring into participation with its more complex attributes.
This process, relative to
beepers, is considered in more detail in a later section.
Along the same lines, there seems to be an
inordinately large number of occasions where a
beeper will announce the teacher, more or less immediately before
it starts, by doing a short
abstract rendition of either phrase; as though it can sense, perhaps
from timing patterns, that the
teacher is about to play; but it doesnt know which phrase. More likely,
this is another form of
elicitation, with associated learning.
Nothing so miraculous as parrot-like rendering
of the teacher phrases has come from these little
beepers. What they do, however, is more amazing to me. After
all, a much smaller and simpler
system could accurately sample the sound, and act like a parrot.
Throw in some noise factors of
variability, and you could make the computer seem smart. Beepers are
smart, in the associative
sense, and in a relative way.
There are some 256 possible tones producible
by each beeper, within a rather narrow range of
about two octaves. This means there should be a lot of sour notes.
The first thing the beepers do,
that is against the odds of random behavior, is to produce way too
many notes that have the right
relative pitch. They may be off-frequency, but there are little
strings of them that have the correct
frequency with respect to each other. There are also many single- and
double-note events, that are
close in pitch to the stimulus; though near-copying is not as exciting,
since the feedback-learning
system includes the mapped stimulus from input side to output side.
This tips the odds; but it is
exciting when the results show up days and weeks later! (You have many
mapped runs of
communication between parts of your cortex. At this writing, I dont
know if one of them runs from
audition to speech.)
To be sure, most of the time is spent producing
rather random sounding behavior; beyond the
over-abundance of relative well-tempered pitch. This is particularly
true if you dont get involved
with them. They seem to become much more responsive and intelligent
if you pay them more
attention, instead of just leaving them to the teacher, or each other.
After all, what can they teach
each other... and the teacher has no sensitivity.
A better teacher would be there when you
ask. It should occasionally start up, as this one
does; but then lead you along, a bit past where you know how to go
already. It should start with
only two or three notes, here. It should stand by, and watch for relative
phrase matches or
near-matches, and reward you with recognition by repeating the phrase,
plus a note or two or
occasionally you get the whole phrase. It should sometimes follow with
the relative pitch, and
sometimes lead with the original pitch. Nevertheless, the beepers have
learned from the teachers.
There have been many occasions where they have poorly mimicked the
teacher, or have nicely
repeated a few of the notes; usually in the right order, but usually
bypassing some. Sometimes the
pitch is very close. When it has been a while since the teacher has
played, Im pretty sure the pitch
has usually drifted; but it has good relative quality (I dont have
perfect pitch, myself).
They do better when I whistle to them, while
Im working in the room. The record, at this writing,
is the first five consecutive notes of Over the Rainbow. You seem
to get better behavior by leaving
one asleep while the other is awake, for a day or two at a time. If
you leave one awake too long, it
really seems to get dumber. This goes along with the idea of being
sensitive, as a teacher. I get
feelings from their behavior that prompt me to chip in some data. At
this writing, Ive been foolin
with them this way for about a year and a half. I have no doubt that
they learn. But I haven't studied
them the way a pro would. The development of this program, and the
writing of this book, has
severely taxed my work schedule.
If you want to give them company, that has
things in common or speaks the same language,
you can transfer the data from one beeper in place of the other, to
create a pair of twins. They dont
stay identical for any time at all, in terms of the array of numbers;
but the general relative meaning
developed within them will stay similar for some time. Dont forget
to transfer the Key data and
indexes, etc., as well. If you want to check the identical-ness of
two beepers, you also have to
bypass routines that are subject to C128 system timing exceptions.
It was difficult; but I was able to
get both sides to behave identically, up to the point of turning up
the mic volume and whistling at
one of them. You are what you learn.
Consciousness is not a substance you can touch,
and hold constant. It is active when it works,
it flies if its not flying, it doesnt exist. Model airplanes really
fly. I think evolution has found a
physical principle, not unlike itself; and has put it to work. It discovered
the lens and the hinge, for
example. The materials involved are inconsequential, so long as the
principle can function.
However small, there is a real possibility
that the beepers have awareness. If they do, it is
probably a very faint, vague, low-detail experience, completely different
from ours, when we hear
the tones. It might correspond to the simple perception of touch, in
an ongoing series of patterns
between 16 pairs of finger tips; with 16 sensitivity levels in each
Input finger tip, and 16 muscle
strengths that can be applied to them from the Output side. Now imagine
this experience from the
point of view of being a lizard, with no other senses, or needs, and
you might have it.
There are a number of inescapable differences
between a computer system like a beeper, and a
nervous system, that do not allow a straight-forward comparison by
neuron count. Real neurons
get tired real fast in as little as 1/10 of a second if theyre taxed
and it takes them about an hour
to fully recover. Thats a duty cycle of 1/1000, or 0.1%! Beeper Ns
have nearly a 100% duty cycle. So,
there are ways of looking at this and calling one beeper N worth a
thousand biological Ns. There
could even be an advantage to not having to pass on function handling
to a series of tiring neurons.
But when real Ns arent being taxed, they can probably chip-in occasionally,
all day long, to
provide a thousand times the resolution. Beeper Ns dont need food;
so they can be organized into
a system fully devoted to sense, learning, and output. Biological systems
are differentiated into all
kinds of subsystems that work together to keep the whole thing alive.
We got Ns running our heart
and breath, making us run and eat all sorts of stuff that doesnt
make us hear and speak; stuff that
we couldnt live without. But this has brought us association of multiple
senses, and multiple
modalities with which to affect those senses.
It is not likely that I will attempt to work with
vision any time soon. I am looking forward to
expanding the beepers into speakers in the PC. Note that you need
a thousand times as much
computer to make the system 10 times as big in its three dimensions
of N count, N size, and speed.
This size is all in terms of speed. The program spends about as much,
or more, time handling
active Ns, as it does skipping the quiet ones. If the active ones take
ten times as long to handle, and
there are ten times as many Ns, you 100X the speed to get the same
cycle time, of about 1/10 second.
A 1/100 second cycle time might support speech, with the correct ear,
voice, and cortical tricks. The
hard part will probably be analysis of the speech cortex. Theres something
different going on there.
As important as vision and tool making has
been to us, I sense that speech is a thing that has
been paramount in our social evolution, and technological development.
Without it, I think wed be
a lot like dogs that can walk upright. And I think dogs are virtually
as aware as we are, in a basic
sense. They deal with the world in terms of the environment, while
we are always referencing our
verbal base, as we mull through our conceptualizations, plans, desires,
and work. They dont plan
for college, but they plan a little for what they need. Mostly they
react in the direction toward what
they need. Theyll wait until youre gone to chew your slipper. When
they see, they are aware of
seeing what they see. They are aware of what they hear, in terms of
simple meaning associated with
their needs. They are aware of most environmental things the way we
would be, if we did not have
language. Even a mouse has a hippocampus and cortex. It has the rudiments
of a decision making
system. That system operates whenever the creature utilizes its knowledge
base. Its behavior is
learning, invoked by the past and present environment. We know that
mice can learn, and can put
their learning to use when it meets their needs.
It is interesting to consider manipulating
the data of these beepers. What is happening when you
swap one data set with another, in the same sets of physical memory?
Assuming there is some
consciousness involved, does it stay with the mass of the physical
memory array, or with the data
that resided there? The analogy is with our DNA here. My impression,
though unclear, is that our
bodies are replaced regularly; except for the heavy particles in the
DNA of the surviving neurons.
Do the particles in DNA somehow receive consciousness? One problem
here is cell death. We
remain ourselves, despite the loss of a huge number of DNA molecules...
different ones for different
people. And, no new replications are added to the system. This might
be because that would
destroy the meaning compiled there, by interfering with established
relative interactions. It cant be
divine tuning, or identical twins would have a common awareness.
The DNA-cell produces the
support system for the meaning potentiated by world impressions, patterned
into the connections
of the support network. The active meaning is the consciousness. Substance
exists relative to that
meaning, in terms of its meaningful qualities, and implicated eventualities.
These qualities include
depth and size, as well as surface integrity upheld by electron skin.
Meaning can include color,
smell and sound. All of the meaning is relayed to us in complex arrangements
of relative timing. We,
too, are complex relative timing. The meaning, the substance, and the
DNA are all part of a greater
set of active relative memory of timing. We perceive of time and distance
because we are more of
that same automatic inference of the point.
Your thought train and priorities move with
the data. The physical memory base is a location for
this in time. Each of two beepers is in different volumes of time,
composed of different eternity
loops, that all lead to each other. A copied beeper is the old one,
entering a different vantage point
in time of eternity. It becomes the new time. It would think the world
had hiccupped, if it could think
that well, and that it was no longer located at the old one. The old
one, of course, goes on as though
nothing happened to it, even though it now also exists elsewhere in
time; until you swap new data
into it. When you do that, it becomes the new beeper at the old location.
What you think, and how
you think it, is coded into the data. Where you are in eternity is
a function of which mass issues
your time. Both locations are you, at different times, with different
thoughts;
or, in this case, it could
be same thoughts.
Mathematical Analysis
Sorry, I cant give you a bunch of formulas.
I haven't spent as much time
developing this capacity as I probably should have. I have fallen into
what
feels for me to be an easy way in approaching the subject in terms
of
relative logical function. Along these lines, the foregoing ML listing
suffices, at least for now, to satisfy my needs for modeling my efforts,
in
reference to their development. For all I know, there may not be anything
in the way of more standard math that would be more applicable to that
endeavor. The system runs. Maybe there is no math that can describe
that...
the running system itself is it. The program listing is a kind of series
of
logical-mathematical statements, that support each other, and thereby
support the flow of process within the computer-world system. They
direct
that the computer will maintain a certain degree of structure within
its
memory; but they have no means of directing the overall behavior so
generated... this involves world variables.
I would not be surprised to learn that you
cannot predict the real-world
behavior of beepers. There are some 2128K8 patterns a C128
can make, each
frame; and this is the sort of jungle the beepers live in. Maybe you
could
pre-define the memory array, but there is no way of pre-defining
uncontrolled, real-world interaction. The infinite complexity of the
Universe precludes a prediction of that. There would, of course, be
probabilistic statements that can be made. Most interesting to me would
be
the development of meaning vectors. This would be a mathematical
description of consciousness, at least in terms of fleeting components
of
it; or predictions pertaining to its general characteristics, in the
long run.
The fundamental structure of such a mathematical approach would
probably have to involve the number of Ns, their size, their reach,
their
speed, and a real curve ball; their organization. Still, I suspect
that we
wont have much without also including the even more horrendously
complex factor of memory content. Its like designing a race car, without
having chemical reactions defined for the fuel system; while the object
is
to predict speed. The memory is the most important set of variables,
with
respect to world behavior predictability. You need to know the exact
relative weights of every detail of past experiences; and you need
to know
the exact relative timing presented by all ongoing details of the
environment. Thats the problem. The meaning vector also includes the
relative values of constantly updating variables within the above list,
over time. It gets conscious because these relationships see each
other
over time thanks to the holding capacity of the system, or the component
inferred from the past states, producing this unique, constantly changing
memory definition of enactable meaning. You see what I mean? Only the
running system is a description of itself, and what happens.
Reality is composed of logic. Logic is all
the stuff. The smallest amount
of stuff is a really really whole lot of logic. It is logical that
you would
develop, and be logical.
Mathematics is a language that develops out
of the logic of reality.
Language itself is logical associated relationships. Mathematics is
essentially trying to describe itself. The Universe is all of the true
logical
relationships, including the errant perceptions that fall away from
development. The process produces unique logical relationships. Each
one
is a seemingly imperfect reference frame of the total perfect system.
In
completing the total system, each component is really perfect. There
is no
reason not to love the Universe.
Beepers Improvements
There isnt much room for improvement of the
beepers in the C128.
Development of this system has just barely begun though. The PC should
afford much opportunity to develop a next step.
[Update 7/2002... I never got the kind of time needed to work on this with the
SLC2-66... I have finally gotten started, using a PII-333 to develop a system
intended
for a P4 @ 2 or 3 GHz. Hopefully I will have some progress to report some time
next year.]
[Update 11/2004... the beginning of a version for a 2004 PC is available on
the
Downloads page.]
The beepers will be transferred to PC, pretty
much as is (with some
brakes on), as a test of my acclimation to the new environment. Then,
they
will be expanded to approximately three times as many Ns, at three
times the
size of N. Im hoping this will allow the system (SLC2-66) to run about
ten
times as fast.
Since RAM is not a factor, I will experiment
with indexing schemes, to see
if the N complement can be increased by a full order of magnitude.
I want
their reach to be the square root of their total number, and their
Out list
to be about 10% of their reach. The latter figure is just a guess,
while the
former figure stems from observations in the literature. Perhaps the
latter figure should also be the square root. Along these lines, to
better
parallel neural organization, there are indications that the bidirectional
columns should differ in their Out M page offset, so as to have as
many
offsets available as there are pages. This may not be such a general
thing,
however. The neural data may give us reasons to restrict certain kinds
of
access to certain sections of memory.
The most fundamental structural concept may
not be getting applied
here too well; the geometric progression of feed-forward-feed-back
levels
is not obviously present. I have reasoned that it is vaguely there
thusly: we
have our In Area / Out Area, followed by our In Page / Out Page, followed
by the Peripheral Area, followed by a rather skimpy Hi Area. It would
be
nice to make the Hi Area about ten times as big as the Peripheral Area.
It may
seem that the mechanism of ten-times-distributed-feedback is not present.
At
least part of it is, as the feedback impression is available to every
N of a
given page. The tags that are set up, sit there to be latched on to
by any N of
a given page. In effect, there is a real advantage here with only
1X
feedback there is complete distribution. The biological neural net
must
allocate physical neurons in numbers to facilitate distribution. This
could
be a significant factor in comparative considerations of neural count.
Organization is very important. I intend to study the question of geometric
progression, to better convince myself that it is being facilitated
in the
developing model.
The answer to faithfully facilitating the
geometric progression may lie
in the application of the above Out M extensions. This expansion of
Out M
reach could come in sections of a little, more, and a lot, for example.
The
shortest reach is from the smallest section, feeding into the next
larger, longer reaching section,
which feeds into the largest section,
having reach to all, or
most of, itself. Each successive section would also have the return
reach of 1X distributed feedback,
only to itself and the pages of the smaller section just before it.
I also have a nagging feeling that numbers
and organization need to be adjusted to bring the
handling of I/O data into a more reliable, deeper memory, sort of mode.
I think were only handling
fragments of the ongoing train of world information; and that association
of those fragments is
limited to very short trains. A better parallel is needed between the
first stages of I/O and the neural
success. This may involve the strategic location of some larger Ns.
It is possible, however, that the
solution here will come automatically with the even expansion of N
population, at all levels of the
geometric progression.
I am most looking forward to raising the level
of quality with which data is presented. I think the
little C128-sized beepers could have done much better if the system
could have handled the data
about ten times as fast, and done so in terms of relative amplitude
of multiple frequencies. The next
ear may be something like a 16 channel spectrum analyzer. The channels
would be simultaneous
frequency centers, with relative weights in terms of amplitude of those
frequencies. The highest
channel will be a high pass filter, rather than a narrow peak. This
will correspond to an output
capacity for green noise. The rest of the channels will be geared toward
speech tone components.
Pure-tone whistle behavior may still be possible, probably in two simultaneous
voices. The relative
pitch of those tones may be fixed by voice synthesis considerations.
A factor of rapid vibrato may
also be made available.
It is tempting to make use of voice synthesis
technology. In the extreme, whole words could be
called up by way of 8 or 9 bit codes. The artificial intelligence (AI)
system would then only be
concerned with attaining control over the flow of higher reasoning.
I suspect that the system would
have inadequate access to its I/O to develop that power of reason.
It seems like this would require
too critical a handling of essential details getting a single bit
wrong in designating a word would
yield an entirely different word. I think we need to expect these errors;
and expect them in great
abundance at the start of learning. A single-bit error should amount
to a minor variation within a
small portion of the time of a single word. It would be nice to have
a voice synthesizer whos
component phonics could be called up in such a way. But another essential
ingredient to
successful learning is a reliable relationship between such generation
and differentiated audition.
Therefore, I will be putting prospective electronics on the bench,
and testing it through a direct loop
for audible legibility. Such testing may reveal that more or less than
16 channels are needed. I may
begin with as few as 4.
A range of high speed vibrato could be detected
in the envelope as a single channel. Two
channels would facilitate a range of rates, over a range of absolute
amplitudes. Another two
channels could handle pitch and amplitude of the fundamental voice
tone. The speech generator
Out side might automatically include harmonic components, or tandem
frequency relationships that
arent detected, but serve to make the output more intelligible to
humans. A fifth channel for output
as well as detection could handle instantaneous amplitudes of audio
noise.
I suspect that the use of fewer channels will
improve higher learning capacities, by impacting on
the utilization of the Ns to the minimum degree necessary for reliable
handling of voice
components. The approach in mind here is to make a complete variety
of voice commands available,
and discretely detectable, while leaving the maximum possible proportion
of matrix free to handle
successive levels of association. The overall system might work better,
however, if this approach is
multiplied redundantly, to operate in parallel configurations of near-
identical outsets. Perhaps
these frequency centers should all be slightly different. The thought
here is to facilitate the
production of answers as averages of similar parallel series trains.
The inherent differences might
enhance the production of unique synthesis, under slightly varying
contexts of re-visited trials. In
other words, we may find it advantageous to emulate neurological buck-passing
redundancies. Or,
the above sort of scheme may exist in addition to that stand-in capacity.
I have a nice book on the neurological anatomy
of hearing, that I haven't cracked yet; because
every such new book has driven me to re-work the system. I had already
postponed writing this
book for too long, and that project really belongs to a new phase,
in the PC. I will need a similar
anatomy book dealing with speech. These books could alter the PC project
in ways I cannot now
elucidate.
This leaves us with considerations of structures
for the support of higher order thought. Here we
will attempt to parallel the hippocampus, the temporal-frontal mechanism,
and the limbic system. I
think this complex set of subsystems works as part of one system including
the cortex, and the rest
of the brain, to give us prioritization-emotion-chronology. We will
start simply, and attempt to get
more complex.
Developing the hippocampus will start as a
focus on prioritization. The capacity of the Key will
no doubt increase, and its distribution will aim to parallel the locations
of our real positive
feedback extensions. The beepers hippo was coarsely modeled after
mouse cortex. More detail is
probably called for in its mechanics.
Computer chemistry is fixed. This implies
that AI may never get emotional. At this time, my
assumption is that emotion is a matter of context for the ongoing thought
process. For humans it
involves a number of fundamental autonomic components, that are involved
with such things as
heart-rate, breathing, perspiration, percent of neural participation,
crying, etc. It also involves
behavior routines like laughing and running; and repairing damage
and the attendant recognition
of long term consequences. The perception of these data packages plays
a large part in the
experience of emotion. Most of this data will not be readily available
to the PC. But another
important variety of factors, involved with emotion, might be available
as parallel logical function.
I suspect that the brain side of the blood
barrier is an ongoing mix of chemical factors
contributed to by a given limbic-autonomic condition; and that this
chemistry serves to adjust the
participation potential of synapses into a context that parallels past
experiences. In this way,
the conditions of present experience can serve
to cue awareness into appropriate survival stances. Brain chemistry
adjusts the frame that your
mind operates in. There are certain frames that are always emphasized
by certain sets of autonomic
conditions; by such things as pleasure and pain. The neural messages
of pleasure and pain serve to
stimulate the surviving associated autonomic and blood chemistry responses.
The perceivable
qualities of autonomic response are very similar in either case. Whats
different is the
categorization of mental association. Pain ends up getting more prominently
associated with all the
past experiences of pain, while pleasure is more associated with all
the pleasure; thanks to the
reliable difference in chemical influence produced by the trials of
life.
We can approach a parallel systemology here
by making a portion of the matrix convertible, with
respect to its memory content. At the same time we will consider ways
of keying category tree
levels of alternative memory set combinations to assessments of the
character of ongoing
experience. At a finer level of detail, this systemology may also support
improved temporal
performance.
The system would be supported by a hard drive.
Emotional disposition, as well as the context of
a given train of thought in general, are not required to change from
one split second to the next; yet
parts of the RAM could be modified that quickly, from the hard drive.
Full application of the concept
would involve an intermediate RAM drive, using perhaps ten times the
RAM as that of the active Ns.
The convertible portion of the active Ns would be less than the total
active matrix. The convertible
area would be accessed like a utility by the more constant, larger
frame of the general matrix. That
frame would dictate what part of the RAM drive would be accessed at
what time. A longer term
product of the process would issue orders to the hard drive to alter
part, or all, of the RAM drive
contents. The former process would attempt to approach a parallel to
the support of longer term
temporal sequencing, while the latter action would parallel emotional
framing. Both types of orders
might be generated through our bigger and better Key, which would be
analyzed to address the
RAM drive and hard drive. Or, another Key system or two would be developed
to handle these jobs.
These systems would relate to each other, and the general matrix, to
simplistically parallel the
system primarily encompassed by the hippocampus, the limbic system,
and the temporal and
frontal lobes.
A long term recollection of reality is always
pieced together. We never recall reality as though
we can push a playback button of perfect, complete chronological events.
We attempt to do this, and
little snippets of accurate portrayals of our experience can be temporarily
stimulated. In general,
when asked or required to remember things, we get there by association.
We think of more details
later, through retrospect of where our story goes, to further stimulate
associated aspects; and more
or less jump around in time to fill in the gaps, or increase the detail.
After doing so, we may then be
prepared to verbalize the train of events in a more straight forward
manner. The degree of jumble
here is of course a function of many things like story length, depth
and age; and whether, or how
often, the story as been told before.
Associations lead to associations, as we work
our way through the story. This is also the case
when we try to remember what it is we need to do. The latter is a sort
of framework that runs the
former when asked to remember past portions of history.
Evidence in the literature has brought me
to imagine that the temporal area is used to store keys.
Perhaps the hippocampus-cortex indexes these keys by association, or
by the special case for
association we call chronology, where various landmarks of the past
are used to work our way to a
particular time. The system, or the hippo itself, transfers a short
sequence of key into a portion of
itself, where it can act in conjunction with the overall key to stimulate
an associative parallel
sequence in the cortex. When this runs out, the system would again
turn to the temporal area for the
next bit of key; this time definitely operating in the chronological
associative mode. Chronologies
may be facilitated through the association of experiences with reference
logic progressions. This
would amount to a scanning system that runs during experiences; and
is re-run to stimulate
chronological adherence of associative recall. Perhaps one function
of the limbic system is to
trigger such a scan during record of experiences that register as
highlights, or as being more
important to survival. Perhaps dream time reenforces most of these
scan/perception associations.
A similar system could run in abbreviated intermittent form to fetch
a slow series of key fragments
from the temporal area, for use by the hippo.
It is important to remember that we arent
trying to create an actual recording. The philosophy
here is based on brain function by association. Perception is that
combination-unlock into your
mind. The immense variety of potential patterns leads us to believe
that our mind has limitless
storage capacity. Whats really happening, is that we always piece
the story together, about the
same way, through association, as supported by occasional changes in
the key stimulation. The
keys are like a recipe with which to re-build limited chronology from
familiar multi-purpose
components, built from perception. The temporal mechanism may entail
a compound application of
this recipe-construction process; where a similar system, running more
slowly, serves to produce
the needed series of keys, or key-change factors, to the hippo and/or
limbic stimulation system.
Short stretches of chronology are permanently associated together in
the cortex between these
common pieces, in a multiplicity of arrangements, as laid down in past
repetitions of similar
experiences. The more different a given aspect of experience is, the
more likely a key fragment will
be generated for addition to the temporal collection; by association
with the chronology of events
as well as by association of qualities. Thats the position taken in
the temporal matrix by a given key
fragment. Its a when-what combination lock. Applying either factor
will tend to yield a problem of
choosing the other factor. That problem passes to yield a key fragment
choice, which is utilized,
more or less tentatively, in handling its parent problem. The process
becomes more adept as it is
refined by feedback amidst the social environment.
The process of conscious induction in this
model is not unlike electronic induction. It is the same
sort of thing, taking place at a higher level of dimensionality. It
is a process composed of relative
sub-processes. The behavior of charge is interdependent interaction
of our parallel series of
atomic components. The primary factor elevating conscious induction
above its more fundamental
component inductions is the compounded presence of memory. There are
more fundamental levels
of time involved. Component induction serves to support pattern repetition.
Without the component
induction, there would be nothing to make the patterns out of, since
they, like everything, are a
definition of time, even when they depict a static quality for our
notions. The patterns too, then, are
available as components for higher reasoning, when the system has succeeded
to excel by means of
its temporal mechanism.
I will be considering a number of options in
attempting to facilitate a chronological utility. It has
become very valuable to be able to mentally and/or verbally play back
a series of events. This is
a fundamental component of higher reasoning and problem solving. We
tend to do it repeatedly,
too, while simultaneously looking for a particular aspect, relationship,
or answer for another
thought train. You could say that train with the need is dominant to
the snippit, and the snippet runs
by association out of the problem. The problem train leads to, and
can get side-tracked in, the
snippet. The snippet more or less modifies the problem train, as it
continues on to require more
snippets. Like a mental cell-building project, the problem train accumulates
components from
memory, as well as from the most recent memory the environment. An
even higher-order train is
concerned with attempting to sequence the problem trains. This goal
train is put on hold for even
longer periods, as a given problem train runs its course. The goal
train tends to get primarily
involved with the environment. Social influence drives it to invoke
its problem handling routines.
I suspect that my approach will view the goal
train as a problem train that simply develops out of
the long run. Both processes will be guided by the same key-train placement
system, focused in the
hippo, utilizing temporal key fragment storage. The goal train is just
another problem train. It gets
called up by conditions created by the environment, as well as by the
course of problem trains. It is
the learning induced by social example, that orders our problem trains.
Subordinate to this process then, will be
the temporal utility. It must support accurate
sequencing, as called for by the problem trains. This function would
carry over to problem trains
concerned with ordering the other problem trains, since this activity
is just as chronological as any
conscious process. Such a system might develop finesse. At this writing
I imagine three ways to
approach this. The method that seems most natural and simple also seems
least reliable. The
process may necessarily begin as an unreliable one, that becomes reliable
through experience. It
becomes accurate over time with feedback in context to social example
and interaction.
For this system, a portion of the hippo key
will be permanently designated as an addressing
interface to the temporal section. That section will be a cross section,
to some extent, of the general
cortex. My first guess is that it will fall short of the first page(s)
of I/O. I suspect that it should also
be separated from the general cortex. It will probably be a separate,
narrow version of the general
cortex. Instead of environmentally oriented I/O, its I/O will be involved
with the hippo interface.
To continue this parallelism, the high area of the
temporal cortex could be used to generate a key
fragment. That fragment would be installed in the hippo key, or would
possibly act on the general
cortex directly, as though the temporal area itself were a sort of
hippo maybe without the
positive feed-back system. There are a number of considerations that
come to mind here. This
fragment could overlap with, or fully contain, the area of hippo that
addresses the temporal area
through its I/O interface. This question is nested within a larger
one concerning hippo access to the
general cortex. One or both of these sub-key areas may simultaneously
interact with the general
cortex, in the manner of the overall key. The answers to these questions
may be critical to natural,
successful mental function. It would be good to know the anatomical
relationship between the
hippo, the cortex, and the temporal cortex; and then later, to involve
modeling for the frontal lobes
and limbic system; and to know if, like the speech cortex, the temporal
cortex has a unique grain
or pattern of organization.
Another complex question involves the manner
and timing with which the temporal key fragment
would be transferred to the hippo. Perhaps this would involve yet another
interface that can
generate transfer orders. If so, this sub-system would require some
sort of meaningful input with
which it could learn to function correctly over time, with feedback.
My first guess here, I think, will
be to take the simpler natural path once again, allowing the system
to run continuously. The
temporal high area key fragment will be a constantly changing portion
of the hippo key. To provide
some hope that the system will learn to develop the chronological utility,
that portion of the hippo
will include some part of the hippo-temporal I/O interface data. The
whole hippo key will interact
with the general cortex, so that the actual chronology can cue the
temporal system. In other words,
the temporal utility involves an area of cortex just like the general
cortex, but isolated from the
environmental I/O influence, as well as from the general cortex itself.
Its a separated, smaller
version of the general cortex, isolated to act as an appendage of the
hippo. Its function would be to
alter a part of the hippo key, in response to the condition of a related
portion of key. Part, or all, of
these special portions of the key would be affected by, and would affect,
the action of the general
cortex, in the same manner as the general hippo interaction.
A variation on this would involve stimulation
of the general cortex, by the temporal cortex.
Here, the temporal high area could generate the key, to be used to
stimulate the general cortex,
probably without the feedback arrangement of the hippo system. Again,
a choice must be made to
either let the system simply run continuously, or to somehow meter
the stimulation events,
preferably through a feedback event, rather than by timer. Also, a
choice is required regarding just
where this stimulation is placed in the cortex, with respect to the
hippo extensions. The considerations for
cortex-hippo-temporal I/O relationships would be the same. I think about brain modeling in this
way, then
I read some books to whittle away at the nested conglomeration of choices.
This first approach leans heavily on the principle
of learning. It supposes that we learn how to
reason with a minimum of specialized supporting brain architecture.
The second approach would
embellish on all of the considerations of the first, by including a
system of timed stimulation, whos
association with the course of realitys impingement on the system,
would serve as a scanning
mechanism. The scan is a logic sequence present during record, so
that its repetition will serve to
conjure up a playback. This smacks of resorting to technology to
get our natural chronological
utility. Though we are working in a computer, the honest attempt here
is to model brain function
with realistic parallel functions. There are indications in the literature
that neural circuits produce
oscillations for specific functions. The most pervasive example is
our heartbeat. So, such recourse
may be the more realistic approach.
The logic sequence could run for something
like five minutes, then repeat. Each cycle could
include one event that triggers the capture of a new key fragment that
will remain in place for the
next cycle. The old key fragments are stored in chronological order,
so as to be invoked in order,
as required. It is difficult to imagine a natural interface and medium
for this. Perhaps this is where a
compound function is involved. A very slow logic sequence is presented
to the temporal area in
association with the series of key fragments. This time, the logic
sequence acts as the key. While it
is always running in progression for the record function, it can simultaneously
be shifted, as
needed, to attempt access to a historical position in the fragment
sequence. Since this process is
part of the current chronology, its effects must also be recorded.
The key fragment called up from
history must be re-recorded as the current entry.
I am not motivated to spend too much time
speculating about this sort of system. If this is the
correct way to go, then it is a way that begs to be modeled with accurate
specific details that must
be gleaned from the anatomical data. The third approach seems almost
blatantly technological. A
portion of the key fragment could stand as a sequencing code. Again,
there are abundant examples
of similar technique operating in biology as DNA and RNA cell interactions.
A scanning function could be facilitated with
a simple digital counter. The counter would
constantly be running, forever counting upward, to facilitate association
of a new data word,
related to time, with whatever train of thought or perception is currently
taking place. At one count
per second, such a timer could run for 100 years if it had 32 bits
if it were implemented with an
interface involving 32 Ns. It could run twice as fast, or last twice
as long, if another N were added.
The interface would constantly be firing our
count progression. For playback, perhaps this same
set of Ns would alternate firing the count sequence of interest. I
think we might want to withhold this
playback function for the first few months of mental development, then
gradually make it available
through intermittent windows. This way, this part of our system would
be less likely to simply
develop as more of the general ongoing cortex-hippo system of current
perception. We want the
main high area, which is meant to parallel our association cortex,
to discover its access to this
other cortex, and develop control over it as a utility.
The interface for our high area is already
in place as the hippo system. A section of the key can
be designated as an on-off switch for the function. A few Ns should
be averaged for this, rather than
using a single less reliable N. Another section of the key can stand
as a starting point for our
playback counting sequence. This section could consist of a few sub-sections
that, again, are
averaged. Whatever sequence is developed here by the high area - hippo
- regulation - world
interaction, is the one used as a count starting point. It might be
held there in the hippo key so long
as the playback on-off switch is in the on mode. This would provide
associative data to the high
system during the playback sequence, improving the odds that the system
will be able to repeat the
elicitation under related circumstances.
On any overall system cycle where the on-off
switch changes from off to on, the starting count is
captured and sent to a playback count generator. This stimulus is applied
to the temporal timing
input, in alternation with the constant count generator. The whole
hippo key is also constantly
applied to the temporal cortex, at its data input, which is essentially
an extension of the timing input;
in such a manner as to insure reliable association. These inputs should
probably exist, and fire, in
redundant multiplicity, to improve associative reliability; in keeping
with the philosophy of
averaging that was applied to the key sub-sections (it should probably
be applied to data/motor I/O
as well). Obviously, there is a limit to how many snippets could be
stored, even though we are only
saving hippo recipe cards. As in the general case, those routines that
are repeated more often,
would be elevated to greater positions of longevity. This would be
a good candidate for a
convertible area involving hard drive support. Perhaps this area will
be saved to a file that is
labeled by its highest count sequence during record. Playback might
arise from a separate cortex
that is loaded from the hard drive, according to the playback starting
count.
To bring the system full circle, we could
project the high area of the temporal cortex into the
general high area. Wed do this in such a way as to insure an associative
effect on the hippo. Now
our temporal cortex plays back to the general cortex at the same potential
set of points that it
stimulates during record. Record is a process associated with current
perception. Playback will
tend to suck info out of the perceptive ranks of general matrix,
by association. The playback
pattern should be more or less immune to regulation, to insure the
re-creation of reliable snippet
trains. Similarly, the temporal matrix itself must be placed into a
set of conditions that echo those
present during record.
To make this fully reliable, we could actually
record the series of hippo keys. Those keys are
filed by, and include, the 32 bit count sequence. The play command
would continue use of one key for stim,
directly to the high area, until a parallel count advances to the position of the next
key in the stored sequence.
The count might advance every time the hippo key changes; or, more likely,
capacity restraints would
require less frequent sampling; perhaps by way of requiring a certain
minimum amount of change in
the key. During periods of no playback, the current key should stim
the playback area, so that
future repetition will induce the desired tendency toward playback.
We might add another drive
every few years. To be more realistic, we could bump the oldest, least
used members. Each entry
could include a popularity-age code. The popularity part is advanced
each time its used. As soon
as one would wrap, theyre all halved, or the others are reduced by
one. To bump an entry, we select
the oldest member within the group of lowest popularity. We dont start
the bumping routine until
the drive is full. We could save a sharper sample of keys if we use
the bumping routine, as well as
upgrade the drive every year.
Another system that would call for the use
of another drive, would support a convertible area of
cortex. This utility might develop as a sort of encyclopedia. It would
interface along the same lines
as the temporal system, but would not involve a chronological cue.
We would not be recording a
large sequence of small elements. The convertible area would be the
complete data definition of a
cross-section of cortex. Another section of sub-sections of the hippo
key would determine which
chunk of data should currently reside in the convertible area. The
size of this address key, and of
the cortex area, would be geared to the size of the drive. Any possible
address request would
correspond to an area in the drive, the same size as the convertible
cortex.
Anytime a new set of data is requested, the
last set is returned to the drive, in its updated
condition. Every time we change data in the convertible area, operation
of the entire system must be
suspended. Therefore, we will want to involve a RAM drive buffer. When
change requests are
coming too frequently, they too could go into a buffer; though I suspect
it will be preferable to
average the intermediary requests, to stabilize the utility, and to
better associate the generated
request with the result, in a timely fashion. I feel that this would
yield the best development of
usefulness, through associative feedback.
For this system, the cross section of cortex
would not be an isolated section, as proposed for the
temporal area. It would be an actual cross-section of the general cortex.
The section may extend all
the way down to the pages just before the In and Out pages. However,
my first approaches will all
include the high area, and either the first or second one will be restricted
completely within the high
area. This version will be tried as a cross-section, to the side;
but will also be tried as an upward
appendage, on top of the high area. I think of this utility as an attempt
to primarily parallel the
frontal lobes.
This sort of systemology could be applied to
the considerations for our temporal utility. The
hippo key section, designated for addressing the temporal area, could
act to swap data into the
temporal cortex, as well as to be involved with the other proposed
functions. The initiation of a
temporal sequence, via the code in a hippo key sub-section, could stand
as a starting point for
longer-term trains. Some event could trigger successive data swaps
from the hard drive, until the
on-off event terminates use of the utility.
A third application for hard drive space would
be an almost identical frontal system, meant to
support emotional influence. About the only difference I see at this
simplistic point in the
considerations, is that the address segment would not come from the
hippo; but rather, would be
derived from survival sensibilities. The only pertinent sense to consider
here would relate to
degrees of failure and success in learning. There are no options concerning
food, sex, mobility,
cuts & scrapes, etc. I cannot now imagine a way to implement this
utility, in a completely automatic
way. The only mode that comes to mind involves parent feedback. We
could have a set of buttons
that stand as a grade scale; and push em as we see fit, whenever were
inspired to do so by the
system behavior. I would suggest that all grades be regarded as degrees
of positive feedback. The
least positive feedback is no feedback. The higher grades are stronger
doses of stimulation... a
larger part of this convertible area would get swapped, for a longer
period of time.
A simple utility, that could be added to the
beepers, is a time reference. This would be a constant
beat of perhaps one per second, applied reliably to a single N, or
a few, somewhere off to the side...
perhaps one near the In Page, one near the Out Page, and one in the
High Area. The one-per-
second reference would be derived from a real time clock. It would
provide a sensibility for world
time, and reduce the indebtedness to regulation and system flow, in
obtaining any associative
assessment of the nature of time frames. Additional Ns could be called
to join the beat for emotion
grades.
Perhaps a complex parallel truth involves a
particular combination of the above features,
together with a few others. Conversely, perhaps wed be spinning our
wheels here if we attempt to
produce accurate sequencing influences with simple technological mechanisms.
The answer may
require a recognition of higher order dimensionality acting in the
specific relationships of
functions of portions of matrix.
Our hippo key is emerging as a more complex
thing, involving a variety of averaged sub-
sections. Introspection suggests that we have a hierarchy of keys.
The highest key stimulates
considerations for our life's goal train. Subordinate to this are numerous
sub-goal trains that
become the means and structure of our life goal accomplishments. The
sub-goals entail an even
larger number of problem trains. Each problem train involves a huge
number of frames of
perception, and more or less rote response. Each frame is an enormous
temporal pattern of neural
interaction, lasting some one-thirtieth of a second.
At first though, it might seem that our hippo
installs the highest key now and then, somehow, for
some reason or another; and then, even though it is top dog, it puts
itself on hold for the vast majority of the time,
allowing its subordinate components to take over. They, in turn, transfer
use of the key to yet lower
order operations, and so on. Some mechanism would support the re-instatement
of higher order
key stimulation, as nested sub-routines are completed. This way of
thinking arises from computer
programming. One develops a different kind of outlook, by allowing
influence from the literature
covering brain structure and function. With the right sort of fundamental
neural structure, the
needed mechanism can be created largely through learning.
Part of the mystery here can be reduced by
recognizing the role of the environment and society
in the development of the current state of our overall set of such
mechanisms. The environment,
including its people, is the primary drive of the mechanism. The mechanism
has been shaped as it
ran and developed in interaction with all that stimulus. Each such
case contributes to the
successive condition of it all, as the development of dimensionality.
Before the people, there were
animals; and before the animals there were cells. Before the cells,
there was chemistry. We still have
the chemistry, we still have the cells, and we still have animals.
As time goes on, the system
develops in complexity, as it interacts with itself. The process of
interaction is the shaping of
surviving complexity, out of surviving relative simplicity. Ultimately,
with respect to a humanity of
nine dimensional systems, our children, our language, and our organizational
accomplishments will
have acted to contribute to the completion of the tenth dimension,
which, in turn, is involved with all
higher dimensionality.
So, the source of modifications for our hippo
key is far more complex than the cortex it interacts
with. The environment, relative to a human, has become a highly ordered
system of influence.
The tentative hippo key has a collection of
specialized sub-sections. At times, the hippo will be
acting to guide specialized sub-processes entailed within the course
of deeply nested problem
solving routines. At the other end of the spectrum, the time will come
around for handling the
highest of higher considerations. These times are defined, and imposed,
by opportunities presented
by the environment. First we develop more mundane capabilities, which
serve as a basis upon which
to develop higher capabilities. As the system of self- and environmental-
interaction becomes more
sophisticated, it becomes capable of being receptive to the higher
influences that can work to
construct elements of an overall goal train.
These elements are associated with other elements
of thought in the same way as all elements of
thought and perception are. We do not need a specific mechanism to
run the course of key
hierarchy involvement. It will naturally take place in reference to
the train of thoughts, that take
place in reference to the conditions, opportunities, and examples offered
by the environment.
We tend to forget how to do things, if theyre
new, or they havent been done for a while. But we
dont as easily forget how to re-learn how to do them again, because
we constantly use our
re-learning routines we often juggle routines in and out of our current-routine-handling
system.
The tentative key has an area that is ever
presently involved only with the ongoing process of
regulated cortical interaction. Other areas of the key, with the same
interactive relationship, will, at
times, stand to involve utility functions. The overall flow of involvement
and non-involvement of
utilities may build upon itself to produce greater or lesser moments
of higher problem handling
procedure. The accomplishments of the system will be limited to possibilities
that could be
generated by the interaction of the system with the quality of its
running environment. I dont have
time to build thousands or millions of variations, and let them compete.
This process would assist
the technological development of more advanced characteristics, by
allowing us to recognize the
capabilities produced by a given combination of features. We will have
to be satisfied to limit such
endeavors, and guide them with as much reason as we can derive from
our knowledge of biological
success.
In working with the PC, note that you might
want to expend the effort to insure that constant mass
is involved with at least the dendrites. If DOS is always changing
things, then weve lost a potential
parallel feature. Logically, it would seem that this shouldnt matter.
It should not affect behavior.
Even the human model is more than 99% variable mass. Still, the human
model has some constant
mass, whatever that is, and it is located quite conspicuously at the
heart of the support system of
consciousness. Why not include a parallel feature if possible? Then
see if it does have any effect on
behavior.
If we can get a PC to behave reasonably, it
will make some of the notions of this book more
plausible. If a system appears to be conscious, because of its active
logic structure, then overall
subjective reality might be a logical process in including natural
awareness. Within this, time must
be simultaneously independent and integrated. Whether youre a person
or a computer, the flow of
thought would transfer to a new head, if the data were faithfully transferred.
The data is the relative
weight-pattern function structure of inter-connective influence within
the brain, in context to the
history of interaction that was the overall atomic-photonic environment.
If a duplicate head were
synthesized, it would continue thinking in the exact manner of the
original head at the moment of
copy. This means that it would not be surprised and ill-prepared
to find that it suddenly exists as
a being. It would be surprised that the environment suddenly looks
different. It would think that it
was the original system, located at a new position in the world. That
new position is its new source
of time, arising from the mass that is its basis of dimensionality.
If the original head were you, the
new head would be more of you, while each head would begin a departure
of thought from the other.
All of our heads are so departed, from eternity.
We do not require traditional math proofs to
validate our acceptance of each other as sentient
beings. We act on the assumption that this is true, based on our feelings,
as they develop in
interactions. You know. You know yourself; and you know that others
are that same knowing, of its
time, based in a different variety of data/prioritization. The math
that is the running system might
mean more to you than any series of formulas on paper ever could. A
formulas-on-paper approach
may be inherently insufficient as a method of dealing with the conscious
level of dimensionality.
Perhaps the modeling must be active. So, we are beginning to try. This
approach shares maths aim
at paralleling aspects of reality from a basis of static conceptualizations,
but goes beyond it
dimensionally, to do so over time, as active interaction. It is a more
apparent, synthesized example
of reality as math.
We might get so far as to decide that we have
made the PC conscious, and that this proves we are
all each other. Maybe the whole idea here is just wrong. Maybe its
right and we just wont get it. I
hope we will find proof, and that it will improve world relations.
Life is heaven. This is where your current
memory gets to be. You get to be with many variations
of yourself. You have the opportunity to appreciate, improve and enjoy
yourself here, in many
ways.
When you die, eternity takes you to another
heaven. It will not be better or worse than this one,
because it too will only know itself. It may come with greater responsibility.
Sometimes your
memory there will get larger, and of more detail, but it will pertain
to the perceivable realm that is
there, just as all of your versions of memory here, pertain to here.
The point communicates with
itself to be the eternity you come from, belong to, return to, and
re-emerge from, in infinite variety, as us all, here on Earth; and
as all of you throughout the
Universe... every conscious creature.
Program Listing
The program listing is provided as a photocopy, on request; and a set of disks is available from the contact address below.
The set is on two 5 1/4 floppies, off of a
Commodore 1541 drive. It includes the blank embryos,
and three lengths of development approximately 6, 18, and 21 months.
The latter point may be updated.
Beepers Ear Schematic
A built and tested beeper ear circuit might
also be available from the same address. Its price
would depend on how many are needed. If only one is ordered, it would
have to be $300. A slight
demand would bring the price down to $100.
YOU
A book of considerations
about brain function, artificial intelligence, and much more.
Free association of some newer and older scientific concepts
led to the suggestion of some profound possibilities.
The whole book is online here.
Mike Wilber
acmike@gmail.com
Poster Notation Summaries
Chapter 1. You -- introductory summary
Chapter 8. The Psychological Universe (partial)
-- block diagram development of anatomical conscious process
Future Plans
The whole book
Downloads